
O que é o Citoesqueleto? Qual é a sua função?
O citoesqueleto estabelece, modifica e mantém a forma das células. Responsável pelo movimento celular e translocação de organelas, cromossomos, vesículas e várias partículas. Seus principais componentes são microtúbulos, filamentos de actina e filamentos intermediários. As proteínas motoras desempenham um papel importante, auxiliando de forma crucial os microtúbulos e os filamentos de actina na motilidade celular e no movimento das partículas intracelulares.
Filamentos de actina
Os menores filamentos do citoesqueleto são os microfilamentos ou filamentos de actina, que possuem funções estruturais, motilidade celular, adesão celular, divisão celular, suporte para estruturas especializadas, contração celular e muscular. Eles podem estar distribuídos por todo o citoplasma, mas geralmente estão concentrados na periferia da célula, próximo à membrana plasmática. Eles são formados a partir de actina, uma proteína globular que possui um sítio de ligação para a molécula de ATP.
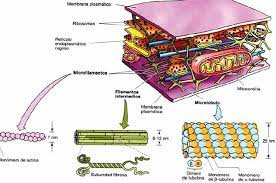
O início da polimerização do filamento de actina ocorre com a ajuda de trímeros de actina, que são formados com a ajuda de proteínas reguladoras, e quando as moléculas de actina são incorporadas ao filamento, é chamado de F-actina. Quando a actina se liga ao filamento, a molécula de ATP é rapidamente hidrolisada em ADP, e as subunidades tendem a se dissociar do filamento. Portanto, se as moléculas de actina forem adicionadas ao filamento mais rapidamente do que a hidrólise do ATP, o filamento crescerá. No caso oposto, o encurtamento do filamento ocorre quando o ATP é hidrolisado mais rapidamente do que novas moléculas são adicionadas.
Dentre as proteínas reguladoras podemos citar aquelas que regulam a formação, como a profilina, que se liga à subunidade de actina, proteínas que regulam os filamentos já formados, como a gelsolina, que se liga às fibras, quebrando-as em menores. o fragmento e a cofilina de ligação à actina é para evitar a adição de nova G-actina.
Os microfilamentos são organizados em células em uma rede ou feixe. As proteínas teciduais de fibrina se ligam aos filamentos de actina na forma de uma rede, mantendo a estrutura da rede. A alfa-actina já liga filamentos de actina paralelos em feixes entre si, e quando estão mais afastados, esses filamentos permitem a interação da miosina (que produz o movimento dos filamentos). Existem também feixes paralelos que estão mais próximos uns dos outros e são importantes para suportar microvilosidades e filopódios. As microvilosidades são expansões da membrana plasmática que aumentam a superfície de contato com o meio extracelular e sua capacidade de absorção de moléculas, e possuem feixes de actina ligados às hifas. Claudicação, filopódios e pseudópodes estão associados à migração celular e são formados por feixes de actina próximos uns dos outros.
As miosinas são as proteínas motoras dos microfilamentos, são uma família de proteínas que possuem cabeças motoras que se ligam ao ATP e possuem regiões de filamentos que interagem com a membrana plasmática ou vesículas. A miosina mais conhecida é a miosina tipo II (atua na contração muscular). No músculo, a miosina se liga para formar fibras de miosina.
Durante a contração muscular, a miosina e a actina se encontram paralelamente para formar um sarcômero, a unidade de contração muscular. Quando há um estímulo de contração, a miosina interage com a actina, fazendo com que o músculo se contraia. Os filamentos de actina sarcoméricos são cercados por troponina e tropomiosina, impedindo que a miosina se ligue aos filamentos de actina. Quando há estímulo para a contração muscular, os níveis de cálcio intracelular aumentam e interagem com a troponina, que então altera a posição da tropomiosina para que a miosina interaja com a actina. O ATP interage com a cabeça da miosina, depois muda de conformação e se liga mais adiante na fibra, hidrolisando o ATP em ADP. A cabeça da miosina novamente se liga ao ATP e puxa a fibra, fazendo com que o sarcômero se contraia.
Os microtúbulos
Os microtúbulos são polímeros proteicos em forma de canudo que não são grandes e existem em uma variedade de conformações, participando de diversas funções celulares, como a formação de cílios e flagelos. São formados por heterodímeros, resultantes do processo de polimerização de duas moléculas de proteína globular denominadas alfa e beta tubulina. As duas proteínas se associam no citosol e formam heterodímeros estáveis que são difíceis de dissociar. Esses dímeros são subunidades de microtúbulos.
Tanto a alfa quanto a beta tubulina têm sítios de ligação ao GTP, mas o GTP presente na alfa tubulina está embutido na estrutura da proteína e nunca é hidrolisado a GDP. Moléculas de GTP ligadas à β-tubulina podem ser hidrolisadas a GDP e desempenham um papel importante no processo de polimerização e despolimerização de microtúbulos.
Nas células, os microtúbulos estão em constante crescimento (polimerização) e encurtamento (despolimerização), e essa constante polimerização/despolimerização é chamada de instabilidade dinâmica. Sequências lineares de subunidades de tubulina chamadas protofilamentos, quando conectadas lateralmente, formam a parede do tubo. Nas células, os microtúbulos são geralmente compostos por 13 protofilamentos.
Os microtúbulos são polarizados porque têm duas extremidades distintas: uma chamada extremidade positiva (+) e a outra chamada extremidade negativa (-). Isso ocorre porque as subunidades de tubulina são sempre adicionadas na mesma direção, com beta tubulina apontando para a extremidade mais e alfa para a extremidade menos. A extremidade positiva tem capacidade de polimerização e despolimerização mais rápida do que a extremidade negativa e é mais energética.
O centrossomo é o centro organizador dos microtúbulos, uma organela não membranosa da qual todos os microtúbulos de uma célula irradiam, pois a agregação espontânea no citoplasma é extremamente difícil de ocorrer.
Algumas proteínas relacionadas são capazes de estabilizar os microtúbulos e prevenir a despolimerização. Os microtúbulos longos também podem ser clivados para produzir filamentos menores, e a proteína acessória responsável por essa função é a catanina, que utiliza a energia gerada pela hidrólise do ATP para clivar os microtúbulos. O corte do filamento altera a dinâmica dos microtúbulos à medida que novas extremidades positivas e negativas são formadas e polimerizadas e despolimerizadas.
As proteínas acessórias que se associam lateralmente aos microtúbulos podem estabilizá-los ou mediar suas interações com outros componentes celulares. Conhecidas como proteínas associadas a microtúbulos, ou MAPs, essas proteínas possuem um domínio de ligação a microtúbulos e outro que pode se ligar a outros microtúbulos ou diferentes componentes celulares.
Outras proteínas acessórias são proteínas motoras, que permitem que os microtúbulos sirvam como vias para o transporte e localização de vesículas e organelas intracelulares e, portanto, são muito importantes.
Devido à hidrólise do ATP, eles mudam sua conformação e, portanto, são capazes de se mover nos microtúbulos, carregando seus ligantes. Dependendo para qual extremidade do microtúbulo elas apontam, essas proteínas se dividem em duas grandes categorias: as cinesinas são proteínas motoras que apontam para a extremidade positiva do microtúbulo, e as dineínas “caminham” em direção à extremidade negativa do microtúbulo. Em geral, podemos dizer que as cinesinas se movem em direção à membrana plasmática e as dineínas se movem em direção ao núcleo.
Ambos os cílios e flagelos são compostos de microtúbulos e dineína e são estruturas móveis.
A proteína motora
O citoesqueleto possui proteínas ligadas a ele, que são proteínas motoras, sensores químico-mecânicos, especializadas em converter energia química em energia mecânica. Assim, a energia da hidrólise do ATP fica disponível para o deslocamento intracitoplasmático, para o qual são formados complexos específicos formados por proteínas e cargas.
As proteínas motoras transportam vesículas, cromossomos, filamentos e até organelas através dos microtúbulos, e fazem parte do sistema de transporte essencial da célula, que consiste em duas partes: adaptadores e motores. O conjunto adaptador é conectado por um lado às partículas a serem transportadas e por outro lado ao segundo conjunto, o conjunto motor. Estes, por sua vez, conectam-se a adaptadores de um lado e ao citoesqueleto do outro, em microtúbulos ou filamentos de actina. As partes móveis fornecem o deslizamento (movimento) dos complexos proteína-partícula ao longo do citoesqueleto.
Uma única célula pode conter vários tipos de proteínas motoras, cada uma especializada para uma carga específica. Essas proteínas podem ser divididas em três grupos: cinesinas, dineínas e miosinas. Independentemente de qual grupo eles pertençam, eles se movem unidirecionalmente ao longo de filamentos ou microtúbulos, de modo que esses componentes do citoesqueleto atuam como orbitais para proteínas motoras.
Duas proteínas motoras estão associadas aos microtúbulos, a dineína e a cinesina. Eles também têm em comum o fato de serem tetrâmeros com cabeças esféricas e ligações de carga-proteína em suas caudas, que podem ser descritas como proteínas que possuem a mesma forma e função, mas não formam filamentos que interagem quimicamente com os microtúbulos. em uma coisa: a direção em que eles se movem. As cinesinas movem-se da extremidade negativa para a extremidade positiva dos microtúbulos e, portanto, tendem a transportar carga para a periferia da célula, mais próxima da membrana plasmática. Em contraste, a dineína se move do positivo para o negativo, trazendo a carga para o centro da célula.
A miosina, por outro lado, está associada aos microfilamentos, formando assim pequenos filamentos e deslocando-se dependendo de sua relação com outros filamentos. As miosinas musculares são do tipo II, são alongadas e consistem em duas cabeças e uma longa cauda torcida que se combina com outras caudas de miosina para formar filamentos grossos, expondo várias cabeças de miosina, cada uma das quais se liga a um ATP e o hidrolisa. A energia dessa hidrólise é usada para direcionar a miosina para a extremidade positiva (+) dos filamentos de actina. No músculo estriado esquelético, a actina está disposta em filamentos finos cercados por filamentos espessos de miosina, permitindo o deslizamento dos filamentos de actina para produzir fortes contrações musculares. Originalmente, as miosinas eram descritas apenas em células musculares, mas hoje sabe-se que podem ser encontradas em diversos tipos celulares, a exemplo da proteína motora miosina V, que transporta cargas como mRNA e vesículas secretoras. célula. – mãe para broto, além disso, ela está envolvida na divisão de organelas entre célula mãe e filha. Portanto, a miosina está envolvida na contração muscular e no transporte de carga.
Notavelmente, as cinesinas estão envolvidas na microreprodução de gametas femininos, na formação de fusos mitóticos e na segregação de cromossomos durante o ciclo celular, enquanto as dineínas estão envolvidas na construção de microtúbulos, além de transportar vesículas. Fusos na mitose e meiose.





























